

|
ćelijska membrana |
|

Aleksandar Ivanovič Oparin
|
Čuveni ruski biohemičar i fiziolog Aleksandar Ivanovič Oparin u svome delu "Poreklo života" (1924) postavlja jedan od osnovnih bioloških principa koji glasi:"U prirodi ne postoji živi organizam , bez obzira koliko jednostavan, koji nije fizički odvojen od svog okruženja". Naime kako se živi organizmi nužno razlikuju od spoljašnje sredine , kako po svojim fizičkim svojstvima, tako i po hemijskom sastavu, oni moraju da poseduju strukturu koja će održavati stalnost unutrašnje sredine. Struktura koja na nivou ćelije ima ključnu ulogu u održavanju stalnosti unutrašnje sredine upravo je ćelijska membrana.
Ćelijska membrana (plazmalema, plazma membrana, plazmina membrana) osim što održava stalnost unutrašnje sredine ćelije:
|
Osim ovih opštih funkcija koje su zajedničke za ćelijske membrane svih ćelija, ćelijske membrane nekih ćelija obavljaju specijalizovane funkcije. Tako na primer membrane nervnih i mišićnih ćelija omogućavaju stvaranje i prostiranje bioelektričnih i akcionih potencijala, dok membrane žlezdanih i pojedine membrane nervnih ćelija omogućavaju izlučivanje signalnih hormona tj. neurotransmitera.
Ćelijska membrana ima ulogu barijere koja razdvaja unutraćelijski prostor od vanćelijske tečnosti, ali istovremeno ona mora omogućiti i selektivan prolazak molekulima i jonima u ćeliju i iz nje.
Jedan od osnovnih postulata biologije i fiziologije jeste da se funkcija zasniva na strukturi, tako da bi izvršila sve uloge plazma membrana mora da sadrži:
- lipide koji membranu čine polupropustljivom barijerom između ćelije i okolne sredine;
- proteine koji su uključeni u transport kroz membranu kao i međućelijske komunikacije i
- ugljene-hidrate, oligosaharide koji su vezani za spoljašnju površinu membrane i omogućavaju ćelijama međusobno prepoznavanje.
|
Ćelijska membrana izgrađena je od lipida i proteina, kojima su pridruženi oligosaharidi. Lipidi i proteini su osnovne gradivne materije membrane i u proseku je količina lipida jednaka količini proteina. Tačnije rečeno, proporcionalni odnos u količini između lipida, proteina i ugljenih-hidrata varira zavisno od tipa ćelije. U membrani tipične ljudske ćelije proteini čine oko 50%, sve vreste lipida oko 40%, dok su preostalih 10% ugljeni-hidrati. Prisustvo oligosaharida u membrani otkriveno je kasnije u odnosu na lipide i proteine. Lipidi i proteini neprekidno klize jedni pored drugih pa grade tečni mozaik .Membrana je mozaik zato što je gradi više vrsta molekula.
Najzastupljenija vrsta lipida u ćelijskoj membrani su fosfolipidi. Struktura fosfolipida bazirana je na molekulu glicerola, čije su dve hidroksilne grupe zamenjene masnim kiselinama, a treća hidroksilna grupa fosfatnom grupom. Fosfatna grupa je polarna , te je taj deo molekula hidrofilan, za razliku od masnih kiselina koje su hidrofobne, te se pružaju na suprotnu stranu u odnosu na fosfatnu grupu.
|
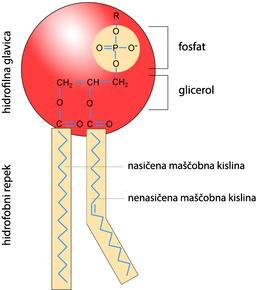
Građa fosfolipida. Hidrofilna glava i hidrofobni rep formiraju izgled slova latiničnog slova V.
|
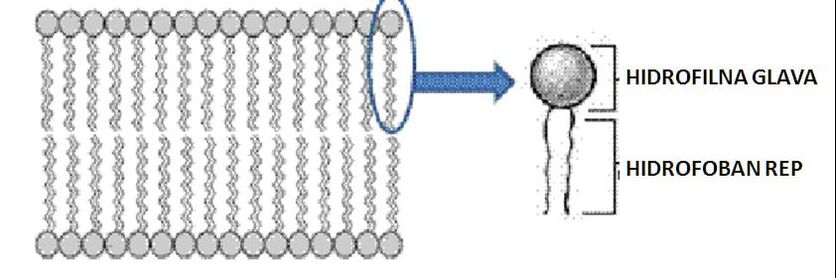
Kako su vanćelijska tečnost i citosol vodeni rastvori sa visokim sadržajem jona, usled hidrofobnih interakcija fosfolipidi ćelijske membrane formiraju dvosloj. Dvosloj fosfolipida se formira na način da su polarne glave okrenute vodenoj sredini, dok su repovi dvosloja okrenuti jedni prema drugima. Fosfolipidni dvosloj predstavlja stabilnu granicu između dva vodena odeljka ćelije. Osim fosfolipida u sastav ćelijske membrane ulaze i:
- Holesterol – koji je ugrađen između fosfolipidnih molekula. On povećava propustljivost lipidnog dvosloja za molekule koji su rastvorljivi u vodi i obezbeđuje stabilnost dvosloja.
- Glikolipidi – koji se nalaze na spoljašnjoj strani lipidnog dvosloja, na samoj površini ćelijske membrane.
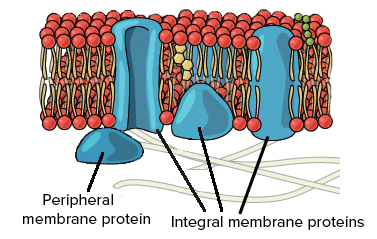
Drugu glavnu komponentu membrana čine proteini.Proteini imaju ulogu u transportu kroz membranu i to su – kanalski proteini koji formiraju kanale za prolazak jona; kao i proteini nosači-pumpe koji prenose molekule iz ćelije ili u ćeliju.
Pored transporta, proteini mogu imati ulogu enzima (učestvuju u reakcijama na površini ćelije) kao i ulogu receptora (za prijem vanćelijskih signala).
Proteini u lipidnom dvosloju mogu biti smešteni na više načina. Proteini koji u potpunosti prolaze kroz lipidni dvosloj, tako da njihov početni i završni kraj ostaje izvan dvosloja, nazivaju se – transmembranski (integralni) proteini. Proteini koji su smešteni samo na spoljašnjoj ili samo na unutrašnjoj strani lipidnog dvosloja nazivaju se – periferni proteini.
Pored transporta, proteini mogu imati ulogu enzima (učestvuju u reakcijama na površini ćelije) kao i ulogu receptora (za prijem vanćelijskih signala).
Proteini u lipidnom dvosloju mogu biti smešteni na više načina. Proteini koji u potpunosti prolaze kroz lipidni dvosloj, tako da njihov početni i završni kraj ostaje izvan dvosloja, nazivaju se – transmembranski (integralni) proteini. Proteini koji su smešteni samo na spoljašnjoj ili samo na unutrašnjoj strani lipidnog dvosloja nazivaju se – periferni proteini.
Osim lipida i proteina u ćelijskoj membrani u manjim količinama su prisutni i ugljeni hidrati. Iako su zastupljeni u malom procentu oni imaju važnu ulogu u procesima ćelijskog prepoznavanja i ćelijske adhezije. Ugljeni hidrati se javljaju najčešće u obliku oligosaharida koji mogi biti kovalentno povezani sa proteinima čineći glikoproteine ili sa lipidima čineći glikolipide.
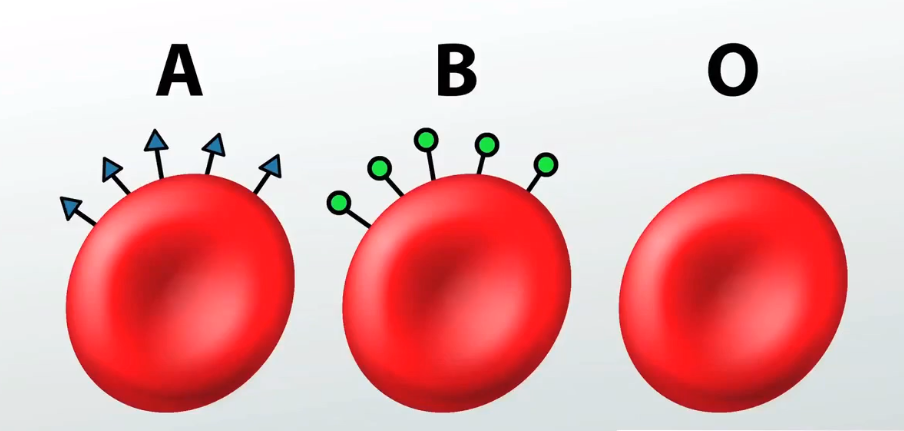
Da li si znao da se po prisustvu određenih oligosaharida na ćelijskoj membrani razlikuju određeni tipovi ćelija iste vrste. Na primer postojanje ABO sistema krvnih grupa kod čoveka, posledica je prisustva različitih oligosaharida u membranama crvenih krvnih zrnaca-eritrocita. Više detalja o ovome možeš pronaći u lekciji o cirkulatornom sistemu. |
|
Jedna od elementarnih funkcija ćelijske membrane je održavanje homeostaze organizma kroz kontrolu supstanci koje mogu da uđu u ćeliju ili da iz nje izađu. To znači da se kroz membranu odvija stalna razmena materije i energije sa spoljašnjom sredinom. Međutim i pored tog intenzivnog saobraćaja , većina supstanci ne prolazi slobodno kroz ćelijsku membranu.Sposobnost membrane da propusti ili ne propusti neke materije je njena permeabilnost (propustljivost). Overton je 1899. g. ispitao više od 500 hemijskih jedinjenja sa gledišta da li mogu da uđu u različite ćelije. Došao je do zaključka da mali molekuli brže prolaze kroz membranu nego krupni molekuli.
Hidrofobni molekuli zbog toga što su rastvorljivi u lipidima lako prolaze kroz fosfolipidni dvosloj membrane. To je slučaj sa molekulima : ugljendioksida, kiseonika i nekih ugljovodonika. Osim hidrofobnih molekula, kroz fosfolipidni dvosloj lako prolaze i neki polarni molekuli pod uslovom da su nenaelektirsani i manjih dimenzija. Tako prolaze molekuli vode i etanola, koji su dovoljno mali da se provuku kroz ćelijsku membranu.
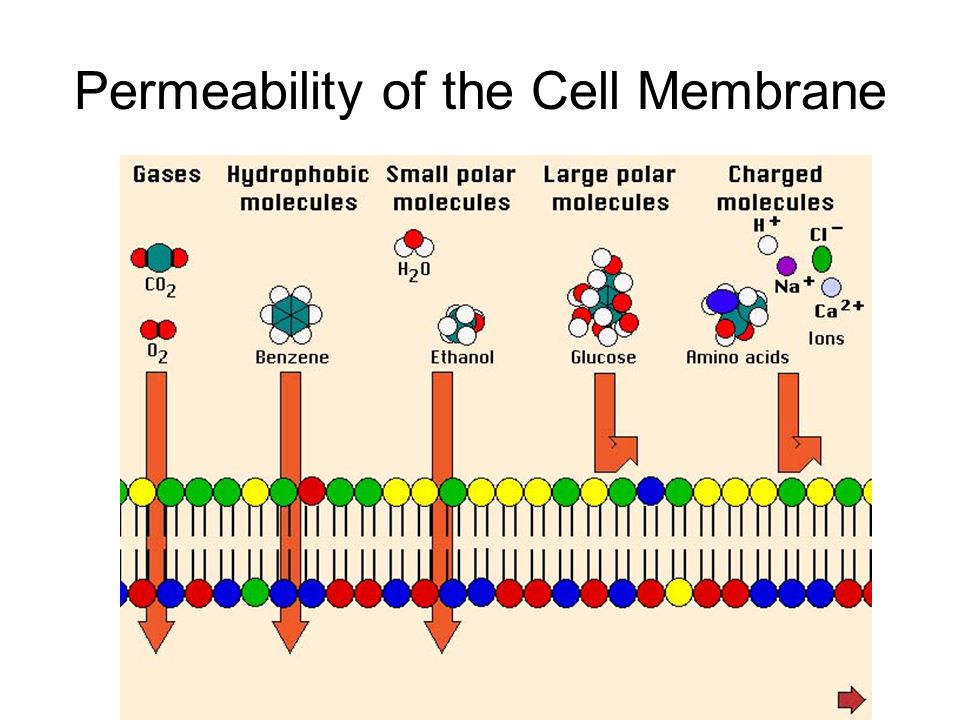
Fosfolipidni dvosloj membrane je nepropustljiv za velike i polarne molekule, kao i za naelektrisane molekule i jone , nezavisno od njihovih dimenzija. Za transport ovakvih molekula i jona zaduženi su transportni proteini ćelijske membrane.
Nezavisno od prirode molekula ili jona koji se transportuje, postoje dva mehanizma membranskog transporta. U zavisnosti da li se obavlja uz utrošak energije ćelijski transport može biti aktivan ili pasivan.
Osnovna karakteristika pasivnog transporta suspstanci kroz ćelijsku membranu je da se on odvija bez utroška energije.
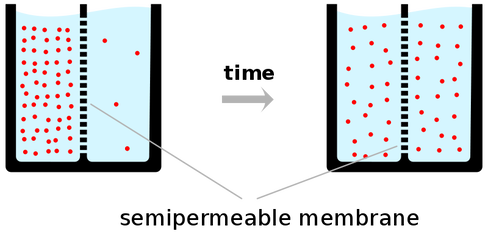 Predpostavimo da polupropustljiva pregrada razdvaja deo suda u kome se nalazi čista od dela suda sa obojenom vodom. Nakon nekog vremena će se celokupna tečnost obojiti, zbog toga što su se molekuli preraspodelili tako da njihova koncentracije bude jednaka sa obe strane membrane.
Predpostavimo da polupropustljiva pregrada razdvaja deo suda u kome se nalazi čista od dela suda sa obojenom vodom. Nakon nekog vremena će se celokupna tečnost obojiti, zbog toga što su se molekuli preraspodelili tako da njihova koncentracije bude jednaka sa obe strane membrane.
Najjednostavniji oblik pasivnog transporta je difuzija. Difuzija je premeštanje molekula iz rastvora veće koncentracije u rastvor niže koncentracije. Prosta difuzija je posledica haotičnog termalnog kretanja molekula. Razlika u koncentraciji molekula između dve strane membrane (u ekstracelularnoj i intracelularnoj sredini) naziva se gradijent koncentracije. Dokle god postoji razlika u koncentraciji molekuli mogu putovati niz gradijent koncentracije bez utroška energije. Postoji prosta i olakšana difuzija.

Prosta difuzija ne zahteva utrošak energije. Za odvijanje proste difuzije potreban je samo gradijent koncentracije molekula koji se transportuje i mogućnost njegovog prolaska kroz membranu. To znači da veliki ili naelektrisani molekuli bez obzira na postojeći gradijent , neće moći slobodno difundovati. Zbog toga je prosta difuzija neselektivan transport, jer ukoliko postoji gradijent koncentracije svaki molekul može difundovati (Izuzev makromolekula i jona). Za molekul koji se prenosi i koji se nakon prolaska kroz fosfolipidni dvosloj rastvori u vodenoj sredini citoplazme kažemo da difunduje. Proces difuzije će se odigravati sve dok se ne postigne dinamička ravnozeža, odnosno dok se ne izjednače koncentracije prenesenog molekula sa obe strane membrane. Prostom difuzijom kroz ćelijsku membranu mogu proći gasovi (kiseonik i ugljen dioksid), voda, etanol i hidrofobni molekuli (supstance rastvorljive u lipidima).
Kao što rastvorene supstance mogu da difunduju, tako i voda može da prolazi kroz membranu sa mesta veće na mesto manje koncentracije. Ovaj proces se naziva osmoza. Osmoza je proces difuzije vode iz hipotoničnog u hipertonični rastvor. Kako je osmoza vid pasivnog transporta (difuzije) ona ne zahteva utrošak energije.
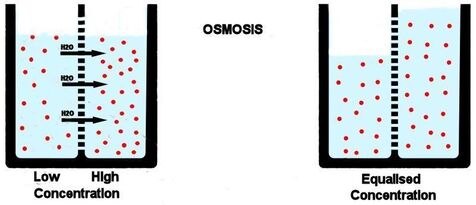
Princip osmoze možemo objasniti na isti način kao i difuzije, pomoću suda sa membranom. Za razliku od prethodnog primera u ovom slučaju uzećemo za obzir da je membrana neporpustljiva za rastvorenu supstancu i da kroz nju jedino može da prolazi voda. Kako je membrana polupropustljiva, sistem teži da uspostavi ravnotežu (ekvalibrijum) između obe strane membrane. Međutim jedini način da se izjednače koncentracije u odeljcima suda jeste taj da voda iz odeljka sa manjom koncentracijom pređe u odeljak sa većom i razblaži je. Za razliku od difuzije, prilikom osmoze se menja zapremina oba odeljka. Zapremina koncentrovanijeg odeljka se povećava ,čime se odeljak razblažuje nasuprot tome zapremina manje koncentrovanog odeljka se smanjuje , čime se odeljak koncentruje.
Za razliku od difuzije koja se odvija niz gradijent koncentracije za jednu supstancu, smer kretanja vode osmozom određen je ukupnom koncentracijom svih rastvorenih supstanci (molekula) u ćeliji. Kada je koncentracija rastvorene supstance izvan ćelije niža nego u citoplazmi, rastvor izvan ćelije je hipotoničan u odnosnu na ćeliju. U ovakvom slučaju voda difunduje u ćeliju dok se ne postigne ekvalibrijum (ravnoteža).Kada je koncentracija rastvorene supstance izvan ćelije veća nego u citoplazmi, rastvor izvan ćelije je hipertoničan u odnosnu na ćeliju. U ovakvom slučaju voda difunduje iz ćeliju dok se ne postigne ekvalibrijum
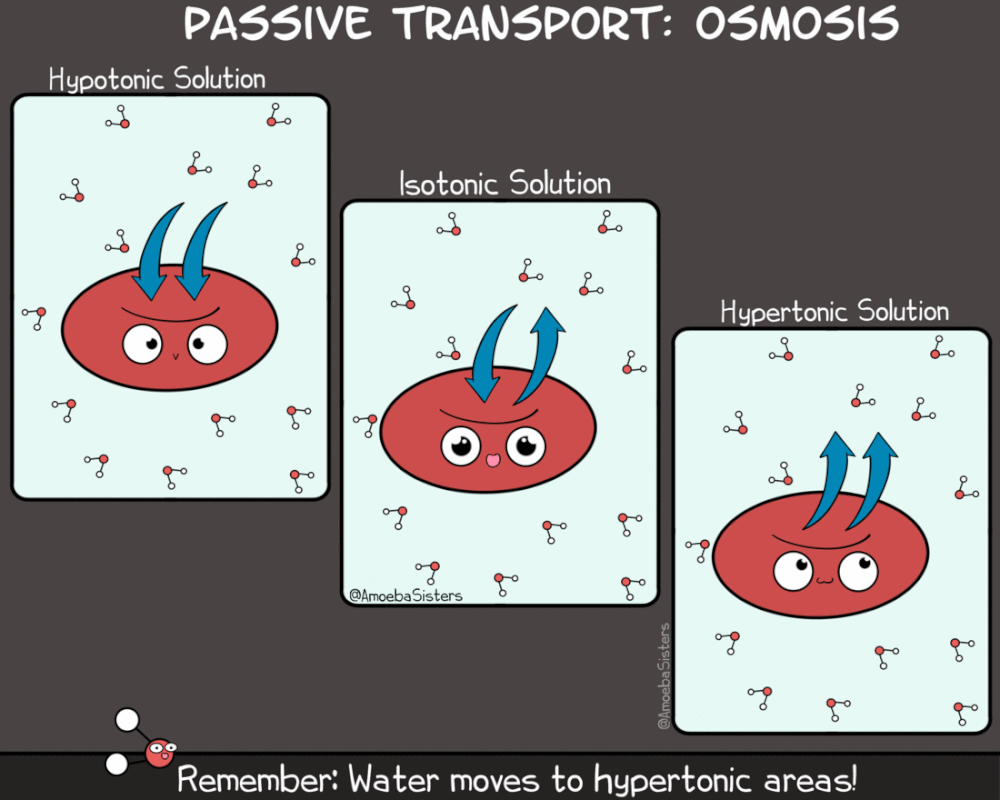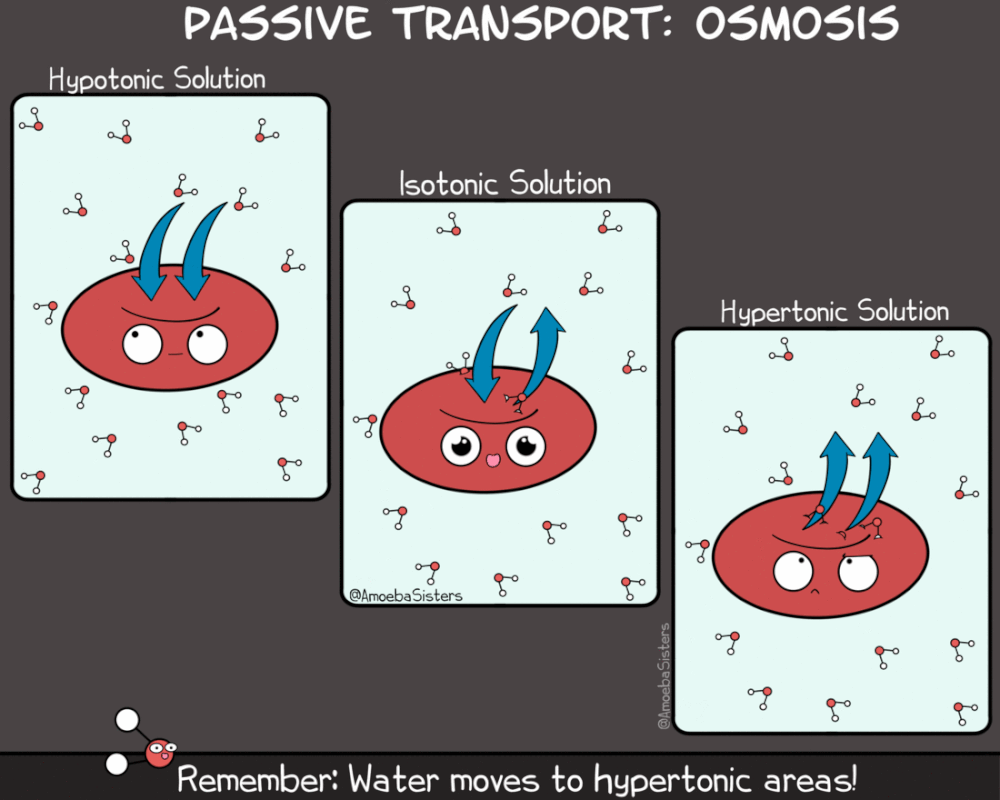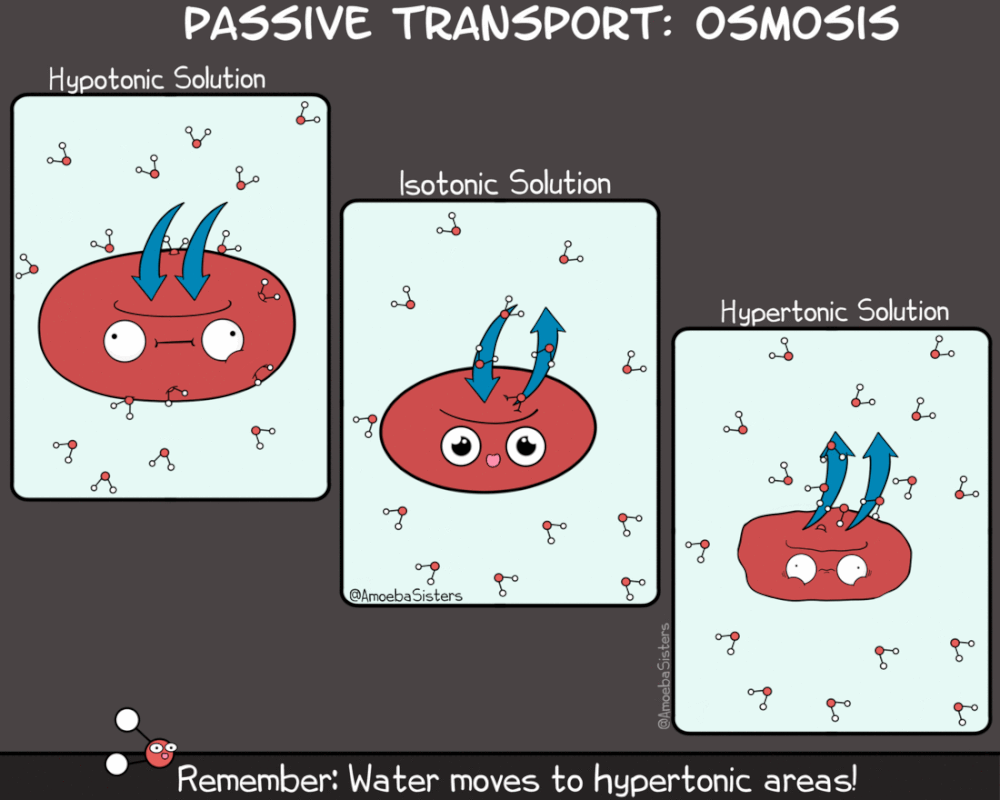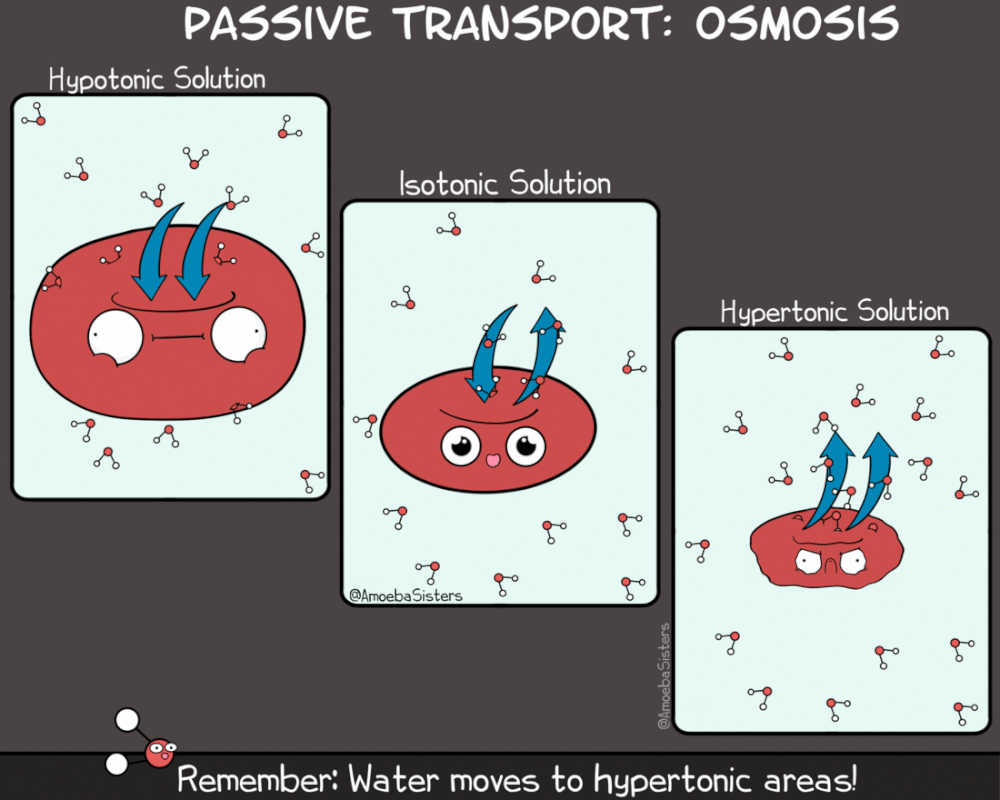
Kada su koncentracije rastvorene supstance izvan ćelije i u ćeliji jednake, rastvor izvan ćelije je izotoničan sa citoplazmom. U izotoničnoj sredini voda podjednako difunduje u ćeliju i iz nje, te se čini da difuzija ne postoji.
Treba napomenuti da se prefiksi hipo- i hiper- , odnose na relativnu koncentraciju rastvorene supstance dva rastvora. Dakle, ukoliko je rastvor izvan ćelije hipotoničan u odnosu na ćeliju, rastvor u citoplazmi je hipertoničan u odnosu na rastvor izvan ćelije . I obrnuto. Voda u bilo kom slučaju ima tendenciju da difunduje iz hipotoničnih rastvora u hipertonične rastvore.
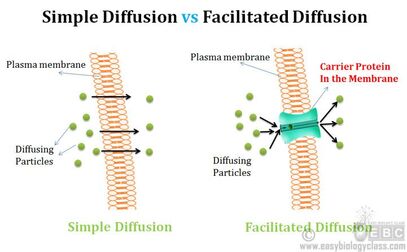 Razlika između proste i olakšane difuzije
Razlika između proste i olakšane difuzije
Olakšana difuzija je proces ulaska u ćeliju onih molekula koji ne mogu da prođu kroz membranu, bez obzira što postoji gradijent koncentracije. Veliki, polarni i naelektrisani molekuli ulaze u ćeliju pomoću posebnih transportnih proteina. Zbog toga se olakšana difuzija naziva još i posredovanim transportom. I prosta difuzija je vrsta pasivnog transporta jer podrazumeva kretanje molekula niz gradijent koncentracije , te se odvija bez utroška energije.
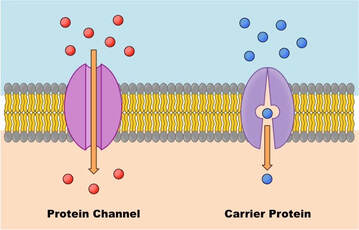
Jedan vid posredovanog transporta odvija se uz pomoć nosača. Nosači su proteinske strukturef u membrani koje vezuju specifične i potom ih prenose kroz membranu u citosol. Po ovom principu u ćeliju se transportuju amino-kiseline i glukoza. Još jedna važna karakteristika olakšane difuzije jeste to da je ona vid selektivnog transporta.Svi transportni proteini membrane su izuzetno selektivni, odnosno olakšanom difuzijom se mogu prenositi samo molekili (eventualno neki od strukturno srodnih molekula) za koje postoje specifični nosači . U praksi to znači, da nosač za glukozu neće moći da transportuje fruktozu ili galaktozu. Osim proteina nosača u ćelijskoj membrani smešteni su i jonski kanali. Difuzija kroz jonske kanale je proces premeštanja jona sa mesta veće na mesto manje koncentracije. Joni i mali polarni molekuli nisu rastvorljivi u lipidima i da bi prošli kroz membranu moraju koristiti prolaze kroz jonske kanale. I jonski kanali su specifični za jedan tip jona, npr. kroz natrijumov kanal mogu samo proći joni natrijuma, ali ne i kalcijuma,hlora itd.
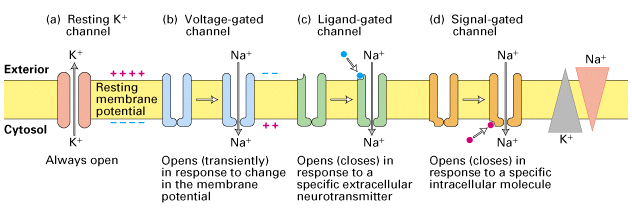
Otvorenost većine kanala zavisi od prisustva odgovarajućeg signala, električnog ili hemijskog. Jonski kanali čija otvorenost zavisi od prisustva električnog signala nazivaju se voltažno-zavisni kanali, dok se kanali koji su regulisani nekim hemijskim molekulom nazivaju ligand-zavisni jonski kanali. Postoje i jonski kanali koji su stalno otvoreni i nazivaju se membranske pore. Osim njih postoje i kanali koji se otvaraju pod pritiskom.
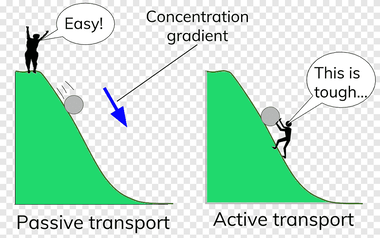 Analogija aktivnog i pasivnog transporta sa guranjem tereta uzbrdo i nizbrdo.
Analogija aktivnog i pasivnog transporta sa guranjem tereta uzbrdo i nizbrdo.
Za razliku od pasivnog transporta, koji se uvek odvija niz gradijent koncentracije određene supstance, transport nekih supstanci kroz membranu odvija se nasuprot koncentracionog gradijenta tj. uz gradijent koncentracije. Ovakav vid transporta zahteva metabolitički rad, a svi procesi u ćelijama koji zahtevaju rad traže i utrošak metabolitičke energije sadržane u molekulu ATP. Ovakav vid transporta kroz membranu koji se odvija uz utrošak energije naziva se aktivni transport. Transportni proteini koji služe za aktivan transport često se nazivaju i membranske pumpe.
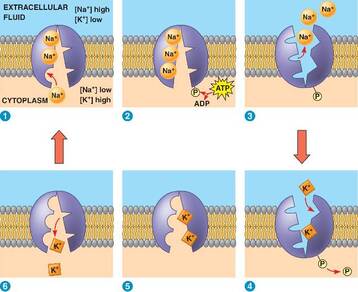
Jedan od sistema aktivnog transporta koji funkcioniše u membranama životinjskih ćelija je natrijum-kalijum pumpa. Kao što joj i samo ime kaže, ova pumpa transportuje jone natrijuma i kalijuma uz njihov gradijent koncentracije. Naime joni kalijuma su uvek u većoj koncentraciji prisutni u citoplazmi nego u spoljašnjoj sredini. Obratno, natrijumovin joni su zastupljeniji izvan ćelije. Nejednak raspored ovih jona uzrok je postojanja razlike potencijala sa dve strane membrane tj. membranskog potencijala što je svojstvo svake žive ćelije. Natrijum-kalijumska pumpa održava ovu razliku u koncentraciji. Tako će natrijum-kalijumska pumpa izbacivati Na+ jone iz ćelije (Iako ih ima više van ćelije), a ubacivati K+ jone (iako su zastupljeniji u ćeliji). Zbog kretanja jona uz gradijent koncentracije troši se energija razgradnjom ATP molekula.
Neke supstance, kao što su makromolekuli i neke veće čestice , ne mogu proći kroz membranu. Zbog toga ćelija koristi endocitozu i egzocitozu da bi takve supstance unosila u ćeliju ili iznosila iz nje. Ovi procesi se koriste takodje i za transportovanje velikih količina malih molekula u isto vreme. Oba ova procesa zahtevaju utrošak energije pa su stoga tipovi aktivnog transporta.
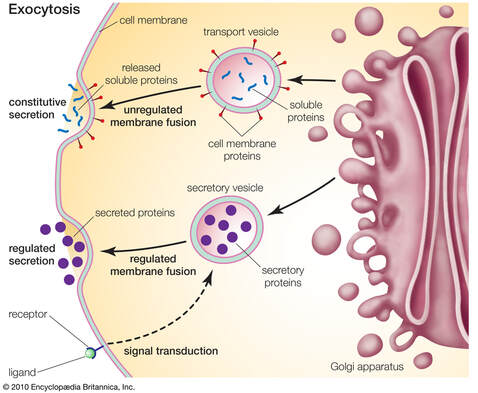
Neki tipovi ćelija kao što su npr. nervne i žlezdane ćelije, izlučuju proteine (ali i druge vrste molekula) koji svoje uloge izvršavaju izvan ćelije u kojoj su nastali. Imajući u vidu da su proteini krupni molekuli , oni ne prolaze slobodno kroz membranu , već se izlučuju uz pomoć transportnih vezikula. U procesu egzocitoze proteini se nakon sinteze i modifikacije pakuju u transportne vezikule. Transportne vezikule putuju do ćelijske membrane , i kada membrane transportne vezikule i ćelijske membrane stupe u kontakt , njihovi fosfolipidni dvosloji se spajaju i sadržaj vezikule se izlučuje izvan ćelije.
Suprotan proces od egzocitoze je endocitoze. Endocitoza je proces preuzimanja molekula ili čvrstih partikula iz vanćelijske sredine , obrazovanjem novih vezikula od ćelijske membrane.Pri endocitozi dolazi prvo do udubljenja na membrani, u koje ulazi materija koja treba da se unese u ćeliju. Udubljenje se sve više spušta u unutrašnjost ćelije, a zatim se ivice membrane spoje i oko unete materije se obrazuje vezikula.
Unošenje krupnih čestica je fagocitoza (grč.phagein = jesti), a rastvorenih materija je pinocitoza (grč. pino = piti). Dok skoro sve vrste ćelija neprekidno unose tečnosti i rastvorene molekule pinocitozom, fagocitozu mogu da vrše samo posebne ćelije – fagocite (npr. leukociti vrše fagocitozu bakterija). Mnogi jednoćelijski organizmi se hrane fagocitozom . Međutim i određene životinjske ćelije-fagociti koriste fagocitozu da bi progutali viruse i bakterije koje napadaju organizam. U tim ćelijama vezikule sa patogenim organizmima spajaju se sa lizozomima, čiji enzimi vrše razgradnju bakterije/virusa pre nego što oni naude organizumu. Endocitoza posredovana receptorom vrši se prepoznavanjem određenog molekula i formiranjem vezikula koja u unutrašnjost ćelije propušta samo te molekule.
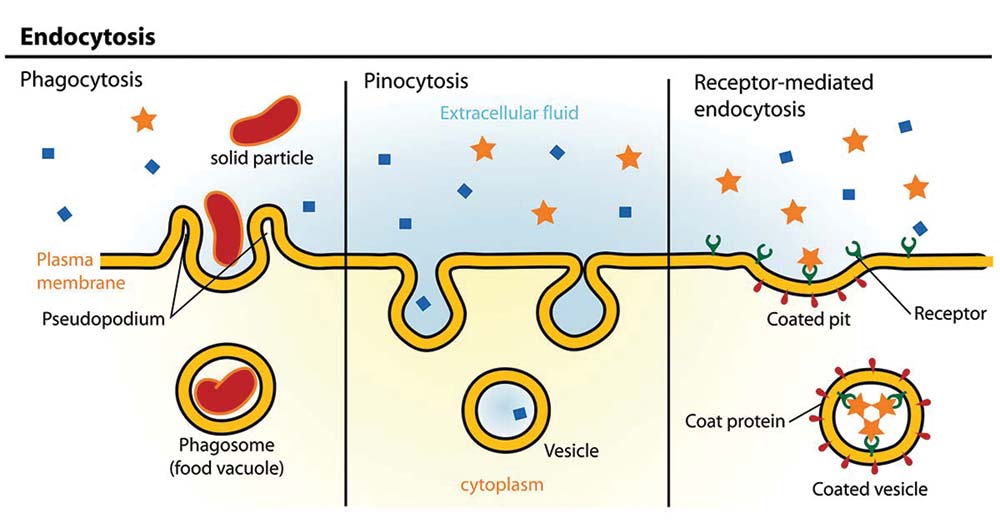
Osnovni tipovi endocitoze
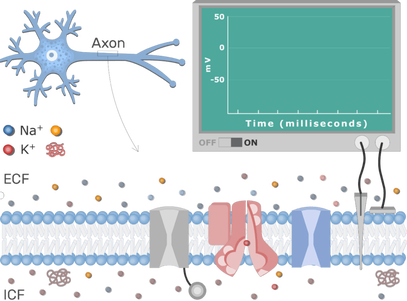 Membranski potencijal se može izmeriti pomoću mikroelektroda i osciloskopa
Membranski potencijal se može izmeriti pomoću mikroelektroda i osciloskopa
Sve žive ćelije odlikuje postojanje razlike potencijala sa dve strane ćelijske membrane tj. membranski potencijal. Ova razlika potencijala iznosi oko -60mV. Prefiks - označava da je unutrašnja strana membrane elektronegativnija u odnosu na spoljašnju. Membranski potencijal nastaje usled nejednakog rasporeda jona sa druge strane membrane, kao i usled selektivne propustljivosti membrane za jone.Koncentracja jona(katjona) kalijuma je intracelularno veoma visoka, dok je ekstracelularno niska. Natrijum je glavni jon (katjon) ekstracelularne sredine, dok ga intracelularno ima samo u niskoj koncentraciji. Najzastupljeniji anjoni u ćeliji su proteini, amino kiseline, dok je hloridni jon glavni ekstracelularni anjon. Ćelijske membrane kod većine ćelija sadrže više jonskih kanala za kalijum u odnosu za natrijum, a kako su jonski kanali veoma selektivni to znači da su membrane propustljivije za kalijum nego za natrijum. Usled veće koncentracije kalijuma u ćeliji javlja se veliki koncentracioni gradijent i joni kalijuma prirodno teže da difunduju iz unutražnjosti ćelije u spoljašnju sredinu. Budući da sa izlaskom svakog katjona kalijuma iz ćelije, ćelijski anjoni ne mogu napustiti ćeliju, ćelija postaje sve više elektronegativnija. U jednom trenutku doći će do stvaranja električnog gradijenta.
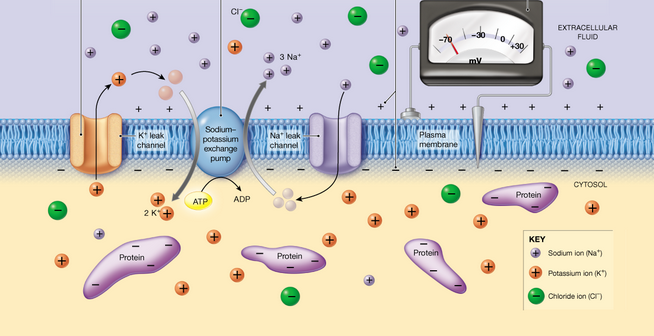
Električni gradijent se suprostavlja koncentracionom gradijentu za kalijum. Pri tome rastuća elektronegativnost unutrašnje strane membrane sve jače privlači nazad jone kalijuma. Joni kalijuma će prestati da napuštaju ćeliju kada se električni gradijent izjednači sa koncentracionim gradijentom. Kod većine ćelija u tom trenutku se postiže potencijalna razlika od oko -75mV. Ovaj napon se naziva ravnotežni potencijal za kalijum. Međutim usled koncentracionog gradijenta, natrijum kroz svoje kanale (koji su oko puta manje zastupljeni) ulazi u ćeliju, te samim tim smanjuje elektronegativnost unutrašnje strane. Zbog toga je vrednost (apsolutna) membranskog potencijala manja od -75mV i iznosi oko -60mV. Međutim ulazak katjona natrijuma u ćeliju, vremenom bi mogao da izazove smanjenje membranskog potencijala daleko ispod ravnotežnog potencijala za kalijum. To bi za posledicu izazvalo preveliki izlazak kalijuma iz ćelije, odnosno izgubio bi se gradijent koncentracije za oba jona. Ovaj hipotetički proces sprečava stalna aktivnost natrijum-kalijum pumpe, koja uz utrošak ATP-a izbacuje 3 jona natrijuma dok upumpava 2 jona kalijuma.. Na ovaj način natrijum-kalijum doprinosi uspostavljanju membranskog potencijala , održavajući njegovu stacionarnost i negativan raspored jona.
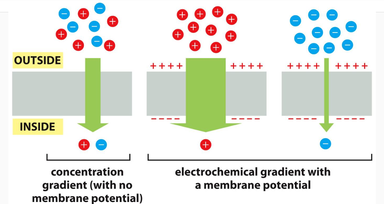 Jon će imati najveću težnju da difunduje u ćeliju kada se istovremeno kreće niz koncentracioni i niz hemijski gradijent (slučaj u sredini).
Jon će imati najveću težnju da difunduje u ćeliju kada se istovremeno kreće niz koncentracioni i niz hemijski gradijent (slučaj u sredini).
Membranski potencijal deluje kao izvor energije od kojeg zavisi transport drugih naelektirsanih supstanci kroz membranu. Zato je od vitalnog značaja održavanje membranskog potencijala. Budući da je unutrašnjost ćelije negativnija od spoljašnjosti , membranski potencijal favorizuje ulazak pozitivnih jona u ćeliju i izlazak negativnih iz nje. Odnosno difuziju jona kroz ćelijsku membranu pokreću gradijent koncentacije i električni gradijent. Ova dva gradijenta (sile) čine elektro-hemijski gradijent. Na osnovu toga možemo zaključiti da se difuzija nekog jona odvija niz elekto-hemijski gradijent.
