

|
gametogeneza
|

|
Gametogeneza je proces obrazovanja polnih ćelija koji prethodi polnom razmnožavanju.Kod životinja krajnji produkt gametogeneze jeste haploidna reproduktivna ćelija,spermatozoid ili jajna ćelija.. U toku gametogeneze dolazi do specifičnih citoloških promena koje uključuju: ćelijsku deobu (dobijanje ćelija sa haploidnim brojem hromozoma – n), jedinstvenu kombinaciju u genomu (hromozomske rekombinacije), ćelijsku specijalizaciju koja podrazumeva da se spermatozoidi i jajne ćelije razlikuju i da imaju svoje specifične osobine.
Razlikujemo dve vrste polnih ćelija-gameta:
- Muški-spermatozoidi
- Ženski-jajne ćelije
U zavisnosti od veličine i izgleda gameti mogu biti:
- Anizogameti-međusobno se razlikuju i morfološki i po ponašanju (po veličini i izgledu). Ovakav vid gameta imaju većina životinja.
- Izogameti- koji su međusobno isti,proces njihovog spajanja naziva se izogamija; prisutni su samo kod malog broja vrsti.
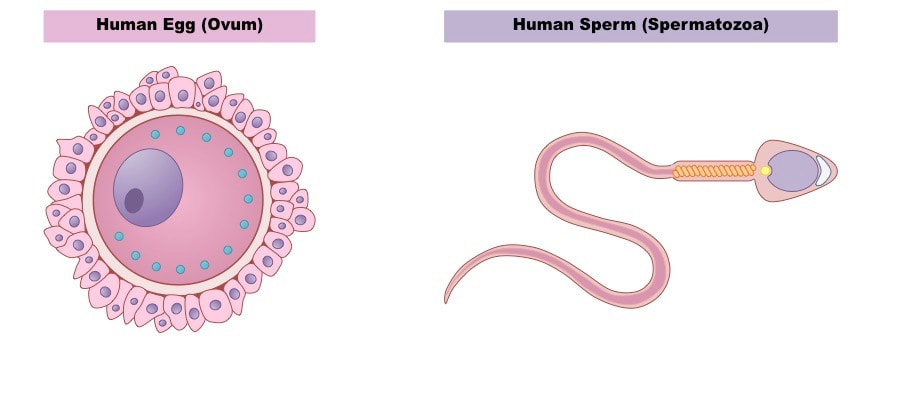
Jajne ćelije su najčešće krupne i nepokretne za razliku od spermatozoida koji su sitni i pokretni.Jajne ćelije(ženski gameti) stvaraju se u ovarijumima(jajnicima), dok se spermatozoidi(muški gameti) stvaraju u testisima(semenicima). Za razliku životinja kod kojih je prisutan polni dimorfizam, kod hermafrodita (dvopolnih životinja-sundjeri,dupljari,puzevi itd. ) u istoj jedinci se stvaraju i muški i ženski gameti.
Iako hermafroditi obrazuju i muške i ženske gamete, redak je slučaj samooplodnje. Razlog je u tome što gameti ne sazrevaju u isto vreme. Na taj način je biološki obezbeđena raznolikost vrsta, tj, sprečena je samooplodnja.
Ćelije od kojih u procesu gametogeneze nastaju gameti nazivaju se primordijalne germinativne ćelije. Ove ćelije nastaju u ranoj fazi embrionalnog razvića i kasnije prelaze u polne organe. Primordijalne germinativne (klicine) ćelije seu muškom polu nazivaju se spermatogonije, a u ženskom oogonije (ovogonije). Od sprematogonija i oogonija mejozom nastaju spermatozoidi, u muškom i jajne ćelije u ženskom polu.
Kod nekih vrsta gametogenetski procesi mogu započeti u embrionu, ali se prekidaju rođenjem i nastavljaju tek kada jedinka postane adult.
Proces formiranja muških polnih ćelija naziva se spermatogeneza, a ženskih – oogeneza ili ovogeneza.
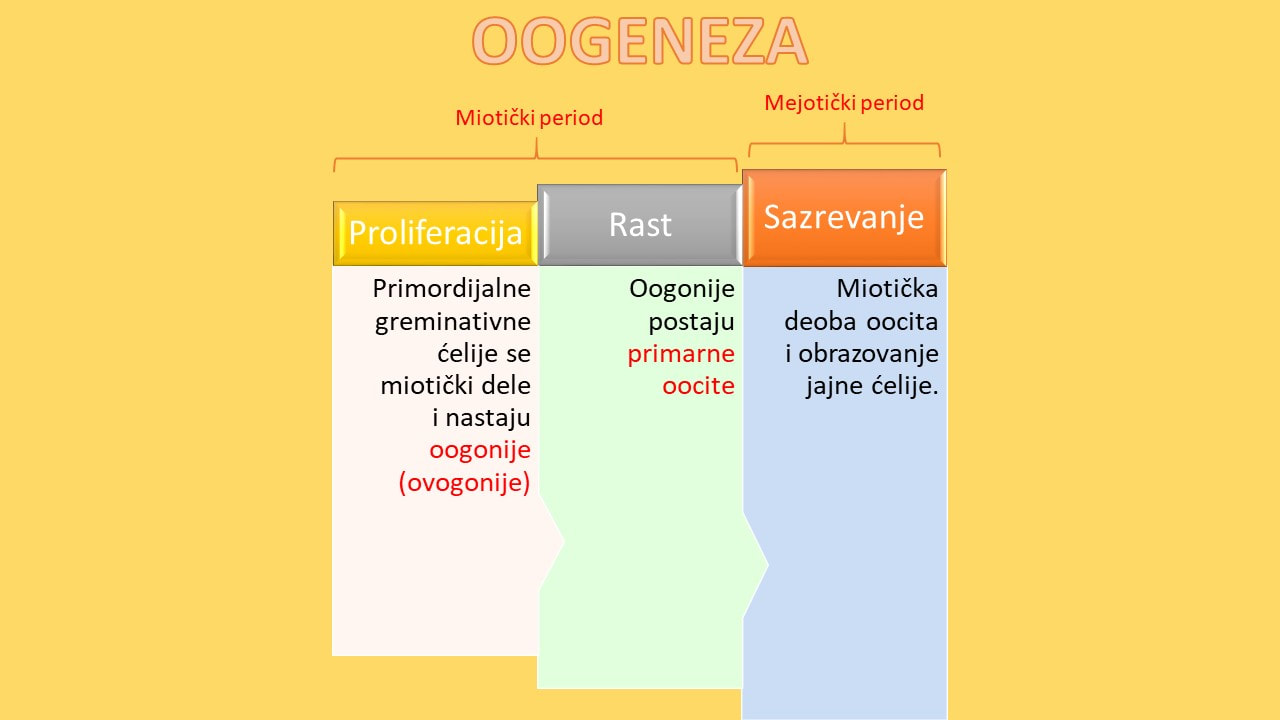
Oogeneza ili ovogeneza je proces nastajanja jajnih ćelija kod diploidnih ženskih organizama mejozom.Mehanizmi oogeneze u životinjskom svetu pokazuju veću varijabilnost i složenost u poređenju sa spermatogenezom. Oo geneza se sastoji iz tri faze:
- Faze proliferacije
- Faze rasta
- Faze sazrevanja.
Faze proliferacije i rasta se mogu svrstati u mitotički period, a faza sazrevanja pripada mejotičkom periodu.
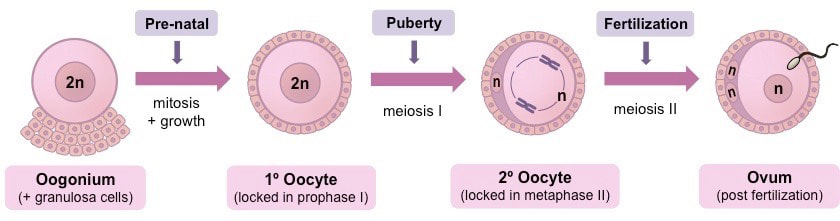
Faza proliferacije započinje još u ranoj fazi ženskog embrionalnog razvića. U fazi proliferacije od prvobitnih klicinih-stem (germinativnih) ćelija mitotičkim deobama nastaju oogonije (ovogonije) sa diploidnim brojem hromozoma.
Sve oogonije ulaze u fazu rasta u kojoj se transformišu se u primarne oocite. Кod većine sisara ovaj proces se dešava s tokom embrionalnog razvića tj. transformacija oogonija u oocite završava se ili pre ili neposredno posle rođenja.
Tako da u jajnicima fetusa starosti od 6 meseci nema greminativnih ćelija – oogonija, jer su sve oogonije diferencirale u primarne oocite. Primarne oocite u jajniku su okružene slojem folikularnih ćelija i nazivaju se primarni folikuli.
Tako da u jajnicima fetusa starosti od 6 meseci nema greminativnih ćelija – oogonija, jer su sve oogonije diferencirale u primarne oocite. Primarne oocite u jajniku su okružene slojem folikularnih ćelija i nazivaju se primarni folikuli.
Starosni uzrast i faze razvića jajne ćelije
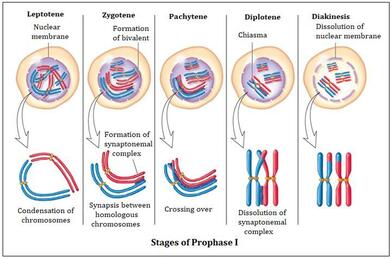 Podfaze profaze mejoze 1. Uoči diploten fazu.
Podfaze profaze mejoze 1. Uoči diploten fazu.
Faza sazrevanja se sastoji od mejotičke deobe. Ovo je ključna faza oogeneze jer u ovoj fazi dolazi do redukcije broja hromozoma na haploidan.
Primarne oocite ulaze u fazu sazrevanja , odnosno u mejozu I još pre rođenja, ali je ne dovršavaju do kraja već se ova deoba zaustavlja u diplotenu profaze mejoze I. To je prvo zaustavljanje i naziva se diploten stadijum (stadijum mirovanja).Za diploten je karakteristično da je to faza u kojoj je upravo završen krosing-over (rekombinacija gena).U ovom stadijumu oocite provode dug period koji iznosi od 10 do 50 godina.
Primarne oocite ulaze u fazu sazrevanja , odnosno u mejozu I još pre rođenja, ali je ne dovršavaju do kraja već se ova deoba zaustavlja u diplotenu profaze mejoze I. To je prvo zaustavljanje i naziva se diploten stadijum (stadijum mirovanja).Za diploten je karakteristično da je to faza u kojoj je upravo završen krosing-over (rekombinacija gena).U ovom stadijumu oocite provode dug period koji iznosi od 10 do 50 godina.
Da li ste zanli da broj oocita vremenom opada?
Broj oocita u oba ovarijuma fetusa starosti 6 meseci iznosi oko 7 miliona. U vreme rođenja ima ih oko 2 miliona. Propadanje se nastavlja i posle rođenja pa ih u vreme puberteta ostane samo 400-500 000
Oocite fazu sazrevanja tj. mejozu nastavljaju tek ulaskom osobe u pubertet (dakle prva oocita miruje oko 10 godina). Počev od prve menstruacije u pubertetu pa do poslednje u klimaksu (kada prestaje reproduktivna sposobnost žene) po jedna oocita u toku svakog meseca završava svoje razviće.
Da li ste znali da starost oocita utiče na njihov kvalitet.
Primarne oocite mogu dugo provesti u stanju mirovanja. Međutim za to vreme one su podložne različitim uticajima i stresrorima.Zbog toga se kod starijih trudnica (preko 35 godina) povećava rizik rađanja dece sa hromozomskim nenormalnostima (hromozomske aberacije) koje uzrokuju teške poremećaje kakav je npr. Daunov sindrom.
Kada organizam dostigne odgovarajuću zrelost, pod dejstvom hormona, primarna oocita tada produžava mejozu I i podeli se na dve ćelije – sekundarnu oocitu i polocitu I- prvo polarno telo.
Sekundarna oocita nastavlja mejozu II-ekvacionu mitozu i zaustavlja se u metafazi II. Tada se nalazi u Grafovom folikulu. Grafov folikul puca čime se sekundarna oocita oslobađa iz jajnika i dospeva u jajovod, što predstavlja ovulaciju. Ćelija zaustavljena u metafazi mejoze II (ootida) predstavlja jajnu ćeliju koja je spremna za oplođenje. Prvo polarno telo (polocita) deli se na 2 polocite.Ako se ootida ne oplodi u roku od 24 časa ona propada,a ako do oplođenja dođe ona dovršava mejozu II, ali tek pošto u nju uđe jedro spermatozoida.
Krajnji rezultat mejoze-oogeneze je da od jedne primarne oocite nastane krupna ootida, koja predstavlja jajnu ćeliju (funkcionalna ćelija) i tri nefunkcionalne polocite.Moglo bi se reći da, za razliku od spermatocite, oocita nikada ne postigne stepen samostalne zrele polne ćelije.
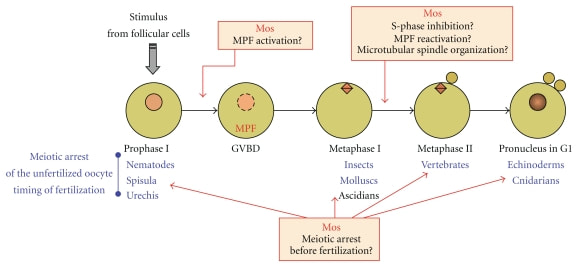
Drugo zaustavljanje mejoze kod različitih životinja. Mejoza nastavlja proces tek nakon oplodnje. Kod kičmenjaka ootida je pre oplođenja zaustavljena u metafazi 2, dok kod bodljokožaca ona u potpunosti završava mejozu.
|
Razmislite o biološkom smislu zaustavljanja oogeneze u mejozi. Uočiti ćete da mejoza nastavlja iz diploten stadijuma tek pošto jedinka dostigne reproduktivnu zrelost. Na ovaj način biološki je kontrolisano, da jedinka može izneti trudnoću, tj. jedinka ne može zatrudneti pre nego što njen organizam nije u stanju da iznese trudnoću. |
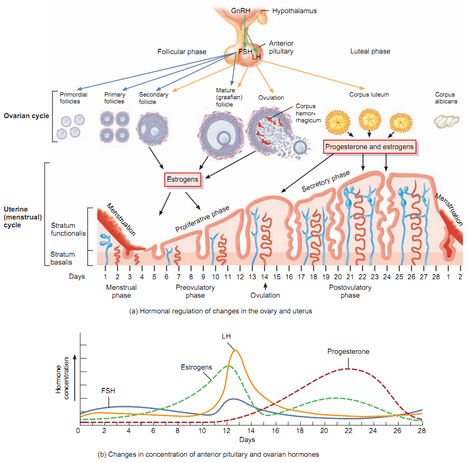 Koncentracije hormona tokom menstrualnog ciklusa i njihova uloga.
Koncentracije hormona tokom menstrualnog ciklusa i njihova uloga.
Proces oogeneze je hormonski regulisan.Prednji režanj hipofize (adenohipofiza) luči hormone :
Osim ovih hormona hipofize, sam jajnik luči hormone koji učestvuju u menstrualnom ciklusu, a to su:
- FSH -folikulostimulirajući hormon i
- LH -luteinizirajući hormon.
Osim ovih hormona hipofize, sam jajnik luči hormone koji učestvuju u menstrualnom ciklusu, a to su:
- estrogen i
- progesteron.
- Estrogen luče folikularne ćelije i uslovljava sazrevanje folikula i ovulaciju. Progesteron luči žuto telo (corpus luteum). Žuto telo se obrazuje posle oovulacije umesto Grafovog folikula. Dejstvom progesterona zid materice se obogaćuje krvnim sudovima čime se priprema za prijem, ishranu i razviće embriona.Ako do oplođenja dođe, onda žuto telo luči progesteron u prvom tromesečju trudnoće kada se obrazuje posteljica koja preuzima funkciju žutog tela, odnosno do kraja trudnoće ona izlučuje progesteron. Ako ne dođe do oplođenja, žuto telo propada čime se smanjuje lučenje progesterona što dovodi do prskanja krvnih sudova materice i menstrualnog krvarenja.
Broj produkovanih jajnih ćelija u jednom ciklusu oogeneze može da varira od jedne(ljudi-žene) do čak pet miliona (ribe).
Iako jajne ćelije svih ♀ životinja nastaju na ovaj način postoji velika raznovrsnost u pogledu izgleda i veličine janih ćelija. Jajne ćelije su najveće ćelije u organizmu. Jajna ćelija žene iznosi oko 0,1 mm, dok jajna ćelija ptica (jaje) može biti veća od 40mm.
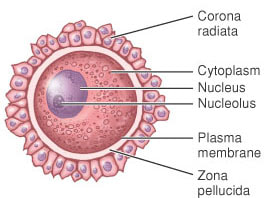
Prema izgledu jajne ćelije su najčešće loptaste ili elipsoidne. Jajna ćelija je okružena vitelinskim omotačem. Vitelinski omotač sisara je pretežno građen od proteina (70%). U njegov sastav ulaze takođe šećeri-heksoze, sijalinska kiselina i sulfati. Kod insekata i riba vitelinski omotač se naziva horion. Membrana jajne ćelije nije ravna i pokrivena je mikroresicama koje zalaze u vitelinski omotač. Jajna ćelija je okružena pomoćnim folikularnim ćelijama. One imaju ulogu da štite jajnu ćeliju , pored toga one luče hormon estrogen i dodatne hranljive materije za jajnu ćeliju. Kod sisara se folikularne ćelije same dele i diferenciraju obrazujući folikul u procesu folikulogeneze .
Jajne ćelije mogu da budu zaštićene i sekundarnim membranama. Sekundarne membrane stvaraju ćelije jajovoda i žlezde koje svoje proizvode izlučuju u jajovod. Ovde spada belance i ljuska jaja kod ptica.
U jajnoj ćeliji se kao osnovna rezervna hrana za embrion nalazi žumance ( lat. vitelus) koje se obrazuje u fazi vitelogeneze prilikom rasta jajne ćelije. Prema količini i rasporedu žumanceta razlikuju se četiri osnovne vrste jajnih ćelija:
- izolecitne-oligolecitne
- mezolectine
- telolecitne
- centrolecitne
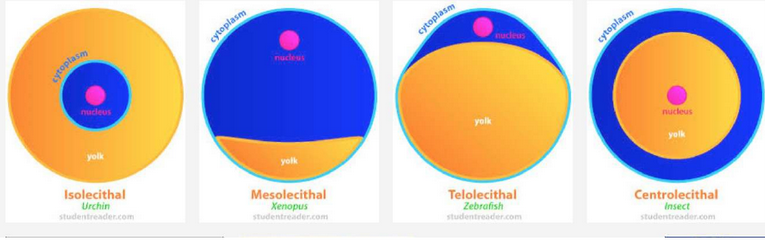
Oligolecitne (oligos= malo; lekhitos = žumance) jajne ćelije imaju malu količinu žumanceta koje je ravnomerno raspoređeno po citoplazmi. Ovakve jajne ćelije imaju bodljokošci i sisari.
Mezolecitne jajne ćelije su srednje bogate žumancetom. Žumance je uglavnom smešteno u jednoj polovini citoplazme. Ovakav raspored žumanceta deli jajnu ćeliju na vegetativnu polovinu (pomoću koje se hrani embrion) i animalnu polovinu (iz koje se razvija embrion). Ovakve jajne ćelije prisutne su kod vodozemaca.
Telolecitne jajne ćelije su izrazito bogate žumancetom. Manja količina citoplazme bez žumanceta je potisnuta na periferiju jajne ćelije. Ovakvu jajnu ćeliju imaju insekti i ptice.
Centrolecitne jajne ćelije imaju veliku količinu žumanceta smeštenog u centralnom delu, dok se citoplazma nalazi oko njega kao površinski sloj. Jedro je smešteno u samom centru žumanceta u jednom ostrvcu citoplazme(npr. jajne ćelije insekata) i povezano je citoplazmatskim mostićima sa površinskom citoplazmom.
Mezolecitne jajne ćelije su srednje bogate žumancetom. Žumance je uglavnom smešteno u jednoj polovini citoplazme. Ovakav raspored žumanceta deli jajnu ćeliju na vegetativnu polovinu (pomoću koje se hrani embrion) i animalnu polovinu (iz koje se razvija embrion). Ovakve jajne ćelije prisutne su kod vodozemaca.
Telolecitne jajne ćelije su izrazito bogate žumancetom. Manja količina citoplazme bez žumanceta je potisnuta na periferiju jajne ćelije. Ovakvu jajnu ćeliju imaju insekti i ptice.
Centrolecitne jajne ćelije imaju veliku količinu žumanceta smeštenog u centralnom delu, dok se citoplazma nalazi oko njega kao površinski sloj. Jedro je smešteno u samom centru žumanceta u jednom ostrvcu citoplazme(npr. jajne ćelije insekata) i povezano je citoplazmatskim mostićima sa površinskom citoplazmom.
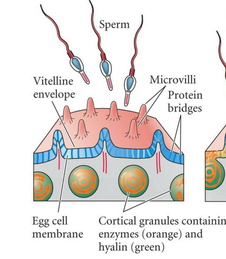
Pored žumanceta od velikog značaja za jajnu ćeliju je deponovanje informacionih molekula razvića. Oni se u citoplazmi jajne ćelije raspoređuju na način karakterističan za svaku vrstu i sadrže informacije o toku razvića. Većina iRNK se aktinskim filamentima transportuje do membrane ćelije i vezuje se za nju posebnim proteinima. Na taj način se na nivou jajne ćelije uspostavlja polarnost. Pored iRNK u jajnoj ćeliji se nalaze brojni ribozomi koji će učestvovati u sintezi proteina. Jajna ćelija takođe stvara i određen broj vezikula- kortikalnih granula. One se nalaze neposredno ispod ćelijske membrane i imaju vrlo važnu ulogu u oplođenju,
Spermatogeneza je proces gametogeneze tokom kojeg nastaju spermatozoidi-muške polne ćelije.Spermatogeneza se vrši u semenim kanalićima muških polnih žlezda (testisa). Za razliku od ženki, muške jedinke se rađaju sa stem (greminativnim) ćelijama u gonadama i zadržavaju ih tokom celog života. Tako da za razliku od oogneze spermatogeneza obuhvata i mitozu greminativnih ćelija (spermagonija) i njihov ulazak u mejozu.
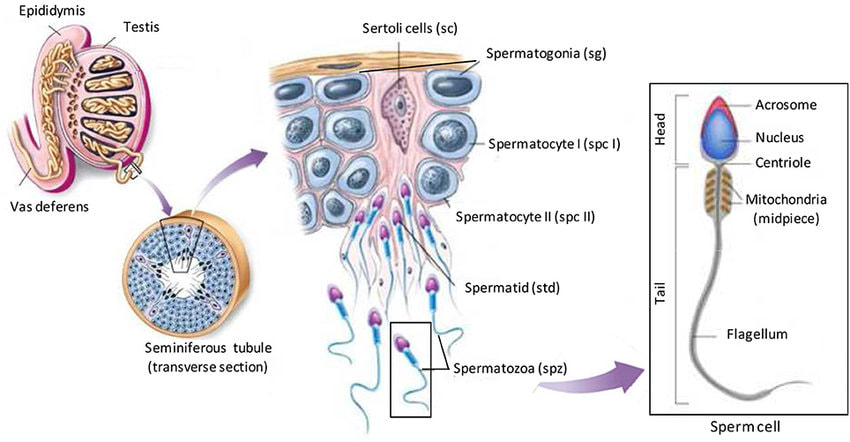
Spermatogeneza se takođe odvija kroz tri faze:
- Faza razmnožavanja (proliferacija)
- Faza rasta
- Faza sazrevanja
U fazi razmnožavanja iz prvobitnih klicinih (germinativnih) ćelija mitotičkim deobama se obrazuju ćelije nazvane spermatogonije (to se vrši još na stupnju embriona). Pošto su germinativne ćelije imale diploidan broj hromozoma - 2n (dve garniture pri čemu jedna potiče od majke, a druga od oca) onda i spermatogonije imaju diploidan broj hromozoma. U semenim kanalićima muškarca nalaze se dve vrste ćelija Sertolijeve ćelije( koje ne učestvuju u razmnožavanju već samo ishranjuju spermatogonije) i spermatogonije.U vreme puberteta među spermatogonijama se mogu razlikovati dva tipa :
- jedan tip produžava da se deli i predstavlja izvor spermatozoida tokom čitavog života (stem-ćelije),
- drugi tip prestaje da se deli i ulazi u naredne faze – fazu rasta i sazrevanja dajući spermatozoide.
U fazu rasta spermatogonije ulaze u pubertetu.Spermatogonije u ovoj fazi uvećavaju svoju zapreminu (rastu) i diferenciraju se u primarne spermatocite. Broj hromozoma se u primarnim spermatocitama ne menja (diploidan ), ali se zato menja količina DNK. U toku faze rasta vrši se replikacija DNK (udvajanje) tako da primarne spermatocite imaju duplo veću količinu DNK u odnosu na spermatogonije.
Sazrevanje se vrši kroz mejozu I i II. U toku faze sazrevanja od jedne diploidne primarne spermatocite mejozom I i II nastaju četiri haploidne spermatide.
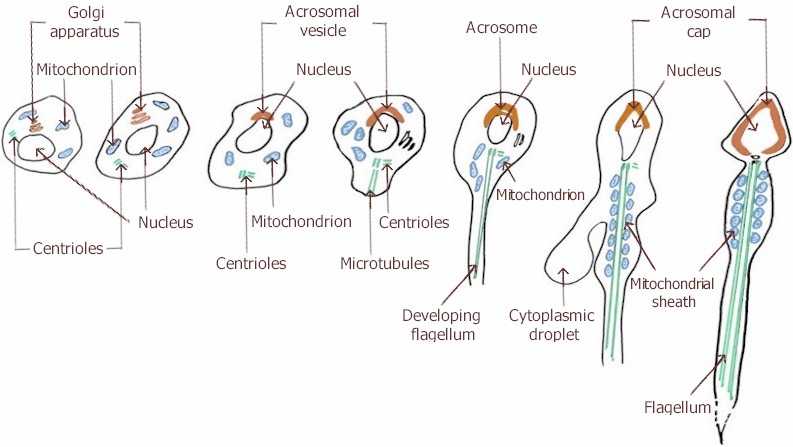
Spermatida, iako haploidna, još uvek nije spermatozoid i da bi to postala ona mora da prođe kroz fazu spermiogeneze (spermatelioze).
Važno:Spermatogeneza i spermiogeneza nisu istovetni procesi! Spermatogenza predstavlja proces kojim se nediferencirane prvobitne klicine (germinativne) ćelija preobražavaju u spermatozoide, dok spermiogenza predstavlja samo deo tog procesa, odnosno, preobražaj spermatida u spermatozoide.
1. obrazovanje biča (repa)
2. formiranje akrozoma ; u akrozomu se sintetišu enzimi koji će pri oplođenju razložiti omotače jajne ćelije
3. odbacivanje najvećeg dela citoplazme da bi se ćelijska masa spermatozoida smanjila i time omogućila njegova dobra pokretljivost
4. kondezacija hromatina da bi se i jedro maksimalno smanjilo.
Ovako diferencirani spermatozoidi se odlažu u pasemenikui još uvek su nesposobni da oplode jajnu ćeliju. Krajnje dozrevanje spermatozoida vrši se u :
2. formiranje akrozoma ; u akrozomu se sintetišu enzimi koji će pri oplođenju razložiti omotače jajne ćelije
3. odbacivanje najvećeg dela citoplazme da bi se ćelijska masa spermatozoida smanjila i time omogućila njegova dobra pokretljivost
4. kondezacija hromatina da bi se i jedro maksimalno smanjilo.
Ovako diferencirani spermatozoidi se odlažu u pasemenikui još uvek su nesposobni da oplode jajnu ćeliju. Krajnje dozrevanje spermatozoida vrši se u :
- epididimisu, gde postaju pokretni i dobijaju zaštitni omotač s kojim nisu sposobni za oplođenje;
- reproduktivnim kanalima ženke (kod žene su to materica i jajovod) gde se zaštitni omotač uklanja procesom koji se naziva kapacitacija (maturacija –sazrevanje).
