

|
FOTOSINTEZA |

|
Život na Zemlji se zasniva na stalnom kruženju materije i energije. Svim živim bićima potreba energija za održanje života.Energija potiče od organskih hemijskih jedinjenja odnosno hrane- gde je energija akumulirana u hemijskim vezama. Energija iz hemijskih veza se može opet osloboditi u procesu oksidacije. Autotrofi u kruženju energije zauzimaju posebno mesto jer su sva organska jedinjenja izgrađena u fotosintezi. Svi ostali heterotrofni organizmi uzimaju gotove organske supstance koje su proizveli autotrofi.

Opstanak svih živih bića zavisi od biljaka. Biljke su jedini organizmi koji su sposobni da energiju sunčevog zračenja pretvore u oblik koji je esencijalan za život. Takođe kao sporedan produkt fotosinteze oslobađa se kiseonik. Za kiseonik ali i za ostala organska jedinjenja smatra se da potiču iz procesa fotosinteze. Skoro svi izvori energije osim nuklearne potiču od fotosinteze.



Fotosinteza je proces pretvaranja svetlosne energije u hemijsku i njeno čuvanje u vidu molekula šećera.Proces fotosinteze se odvija u biljkama, te u nekim bakterijama i algama, koje u svojim ćelijama imaju hlorofil. Za proces fotosinteze biljkama je neophodan ugljen-dioksid, voda i svetlosna energija. Proces se odvija u hloroplastima, malim organelama u citoplazmi biljnih ćelija koje sadrže zeleni pigment zvani hlorofil. Fotosinteza se najvećim delom odvija u listovima biljaka, a veoma malo ili nikako u drugim delovima biljke (stablu, korenu i dr).
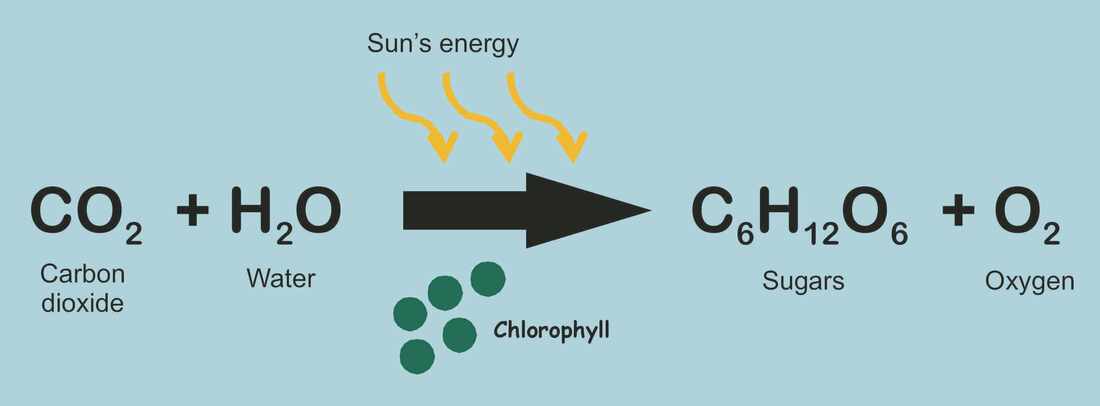
Zbirna jednacina fotosinteze
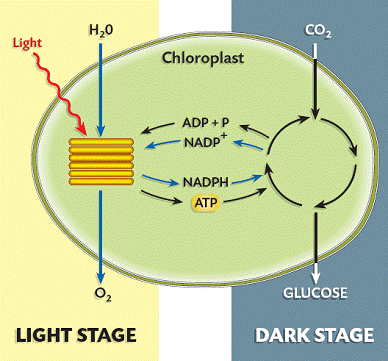
Fotosinteza je veoma složen proces u kome se odvijaju desetine biohemijskih reakcija. Fotosinteza se odvija u dve faze: fotohemijsku odnosno svetlu fazu i tamnu fazu poznatiju kao kalvinov ciklus.Fotosinteza se odvija u dve faze. U prvoj fazi, u reakcijama koje zavise od svetlosti ili svetlosnim reakcijama hvata se energija sunca (ili drugog izvora svetlosti) i koristi u pravljenju molekula u kojima se skladišti energija: ATP i NADPH. U ovoj fazi dolazi i do proizvodnje kiseonika koji se oslobađa u atmosferu .U toku druge faze, u reakcijama koje ne zavise od svetlosti koriste se ovi proizvodi da se uhvati i redukuje ugljen-dioksid.Većina organizama u kojima se vrši fotosinteza i ispušta kiseonik koristi vidljivi deo spektra za fotosintezu, ali su otkrivena najmanje tri organizma koji koriste infracrvenu svetlost.
Kada svetlost padne na neki pigment ona može da bude reflektovana,propuštena ili da bude apsorbovana. Samo ona svetlost koja je apsorbovana je aktivna u nekom biološkom procesu. Od celokupne elektromagnetne radijacije Sunca tokom fotosinteze se apsorbuju samo određene talasne dužine vidljivog dela spektra elektromagnetnog zračenja.
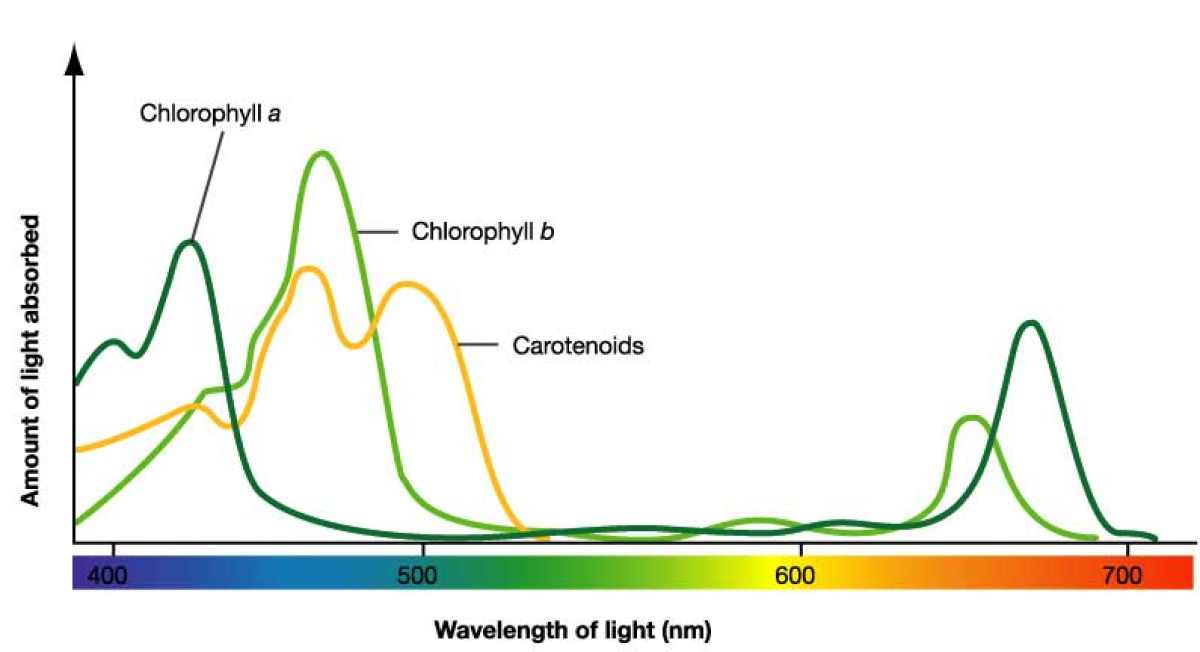
Svetlost koja je biološki aktivna u procesu fotosinteze tj. koju biljke apsorbuju je svetlost talasne dužine između 630-760 nanometara i svetlost talasne dužine između 430-490 nanometara. Odnosno u fotosintezi je aktivna crvena i plava svetlost, što se može videti sa grafika. Pigmenti koji učestvuju u fotosintezi mogu biti samo oni koji apsorbuju crvenu i plavu svetlost.
|
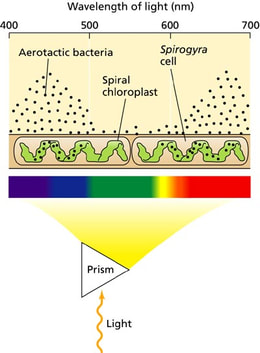
Teodor Vilhem Engelman je bio nemački prirodnjak koji je još pre 120 godina odredio apsorbcioni spektar fotosintetičkih pigmenata. Polazeći od činjenice da se u fotosintezi oslobađa kiseonik on je posmatrao pod mikroskopom deo alge Spirogyra sa bakterijama truljenja. Kako su bakterije truljenja aerobne one se kreću ka mestima gde je veća koncentracija kiseonika.Kada se mikroskop osvetli belom svetlošću bakterije se okupljaju okom dela ćelije gde se nalazi hloroplast jer on izlučuje kiseonik. Engelman je razložio belu svetlost pomoću prizme i osvetlio samo jedan deo hloroplasta crvenom,zelenom i plavom svetlošću. Bakterije truljenja su se okupljale samo oko onih delova hloroplasta koji su bili osvetljeni crvenom ili plavom svetlošću . |
U fotosintezi učestvuju 3 grupe pigmenata hlorofili, karotenoidi i fikobilini. Zbog svojih specifičnih osobina jedini od navedenih pigmenata koji je sposoban da otpočne proces fotosinteze je hlorofil. Ostali pigmenti imaju samo pomoćnu, ali ipak važnu funkciju.
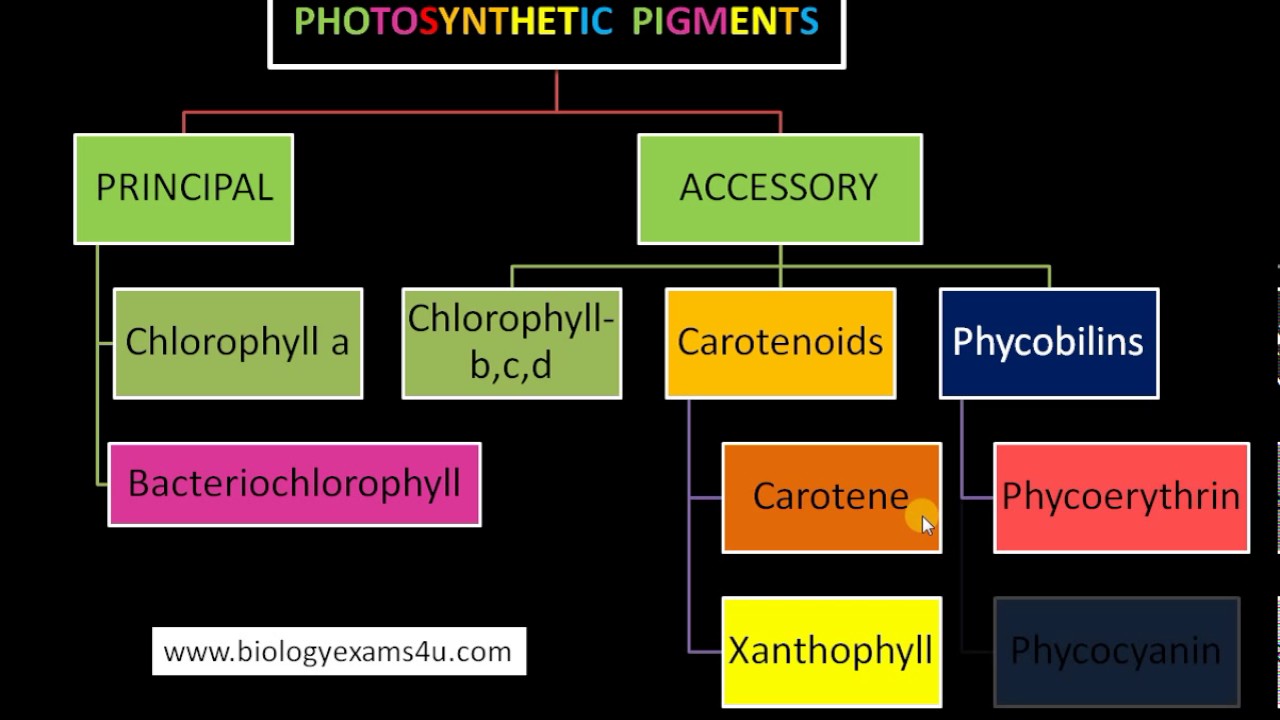
Podela fotosintetičkih pigmenata na glavne i pomoćne
Postoji više vrsta hlorofila. Hlorofil a sadrže svi organizmi koji vrše fotosintezu.Više biljke sadrže hlorofil a i b ; a razne grupe algi pored hlorofila a sadrže hlorofile d ili c.

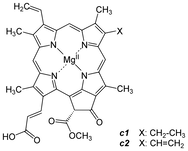
Hlorofil je složen molekul , sastoji se od porfirinskog prstena u čijem je centru atom magnezijuma i alkohola metanola i fitola koji su za prsten vezani estarskim vezama. Karotenoidi su pomoćni pigmenti u fotosintezi. U grupu karotenoida spadaju karoteni i ksantofili. Fikobilini se nalaze kod prokariota. Fikocijanin plavozelene boje se nalazi kod bakterija iz grupe cijanobakterija, dok se fikoeritrin nalazi kod crvenih algi (rhodophyta).Kod fotosintetičkih bakterija takođe postoji specijalna vrsta hlorofila-bakteriohlorofil.
Hlorofili imaju 2 zone apsorpcije u plavom i crvenom delu spektra. Zelenu boju hlorofili ne absorbuju već je propuštaju odnosno reflektuju i zato su zelene boje.Karotenoidi apsorbuju svetlost u ljubičastom i plavom delu spektra, dok fikobilini apsorbuju zelenu,žutu i narandžastu svetlost.
Pomoćni pigmenti povećavaju efikasnost fotosinteze tako što apsorbuju talasne dužine koje hlorofil propušta., a apsorbovanu energiju prenose na hlorofil koji je koristi za dalji tok fotosinteze. Autotrofni organizmi su adaptirali na okolinu svetlost tako što stvaraju pigmente čija je boja komplementarana boji raspoložive svetlosti. Na primer u površinskom sloju mora žive zelene alge koje apsorbuju plavi i crveni deo spektra. Na većim dubinama žive crvene alge one apsorbuju zelenu,žutu i narandžastu svetlost koju ne apsorbuju zelene alge i koja prodire do tih dubina.Količinski odnos hlorofila a i b u telu viših biljaka zavisi od količine svetla kojom su izložene. Tako dobro osvetljene biljke imaju tri puta više hlorofila a nego hlorofila b, dok one koje žive u senci imaju više hlorofila b. Uzrok ove pojave je takođe komplementara hromatična adaptacija koja se manifestuje i na višim biljkama.

Komplementarna hromatična adaptacija. Algama kojima je dostupna zelena svetlost su crvene boje, a algama kojima je dostupna crvena svetlost su zelene.
Hloroplasti su ćelijske organele svojstvene za organizme koji vrše fotosintezu. U njima su smešteni fotosintetički pigmenti. Hloroplasti su obavijeni dvema membranama, a unutra hlorplasta nalazi se matrix sa tilakoidima.Hloroplasti različitih biljnih vrsta se razlikuju po obliku veličini i broju. Hloroplasti se najviše nalaze u ćelijama listova, ali su prisutni u ćelijama svih zelenih delova biljke. Hloroplasti algi se razlikuju po formi, broju i veličini. Neke alge poseduju samo jedan hloroplast koji je znatno veći od biljnog i naziva se hromatofor.

različiti oblici hromatofora
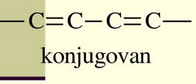
Organske molekule kao što je hlorofil karakteriše poseban tip elektrona koji ne pripadaju ni jednom atomskom jezgru ,već su zajednički za dva i grade vezu između njih.Oni se nalaze u spoljasnjem elektronskom omotaču molekula, ovi elektroni su značajni jer su oni zaduženi za apsorpciju fotona. Ovaj tip elektrona je karakterističan za organska jedinjenja sa konjugovanim vezama. Broj konjugovanih veza direktno je proporcijalan talasnoj dužini svetlosti koja se apsorbuje, tj što je broj konjugovanih veza veći to je talasna dužina svetlosti koja se apsorbuje veća.
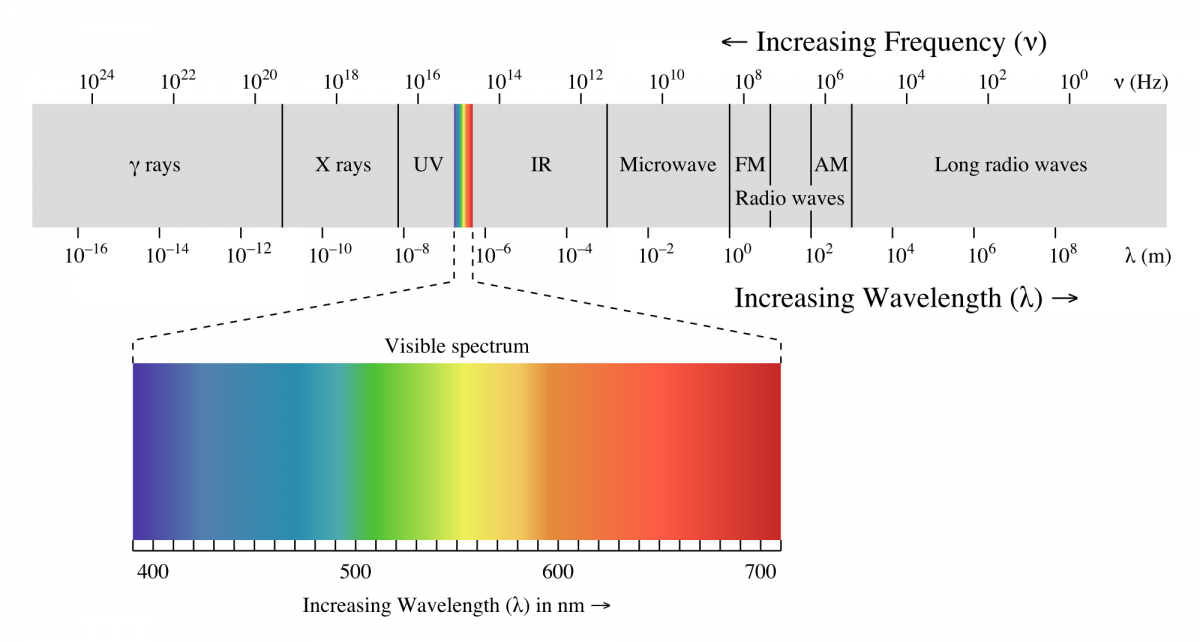
Sa porastom talasne duzine opada energija koju nosi foton
Ukoliko jedinjenje ima više od 7 konjugovanih veza onda apsorbuje fotone iz vidljivog dela spektra elektromagnetnog zracenja. Kako svi fotosintetički pigmenti imaju više od 7 konjugovanih veza oni apsorbuju svetlost iz vidljivog dela spektra, a ne kratkotalasne .
Elektroni koji čine spoljašnji omotač molekula vezani su za svoje orbite i na normalnoj biološkoj temperaturi nalaze se na najnižem energetskom nivou-u osnovnom stanju. Kada se kvant svetlosti odnosno foton apsorbuje elektron prelazi u ekscitirano odnosno pobuđeno stanje. Hlorofil ima 2 orbite pobuđenog stanja višu koja odgovara svetlosti plave boje i nižu koja odgovara svetlosti crvene boje. Ekscitirano stanje molekula je vrlo nestabilno i kratkotrajno. Do oslobađanja te energije (deekscitacije) dolazi na više načina i to veoma brzo. Kada do deekscitacije dolazi usled gubitka elektrona počinje fotosinteza.
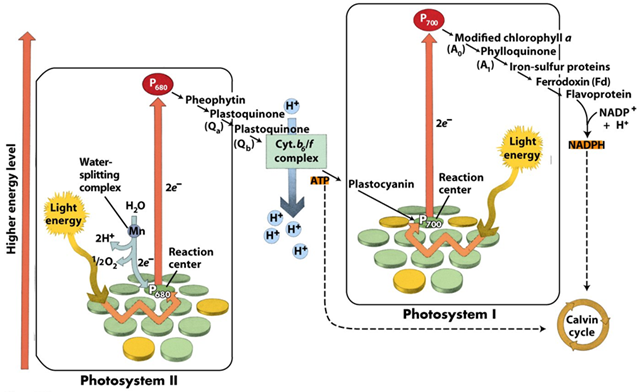
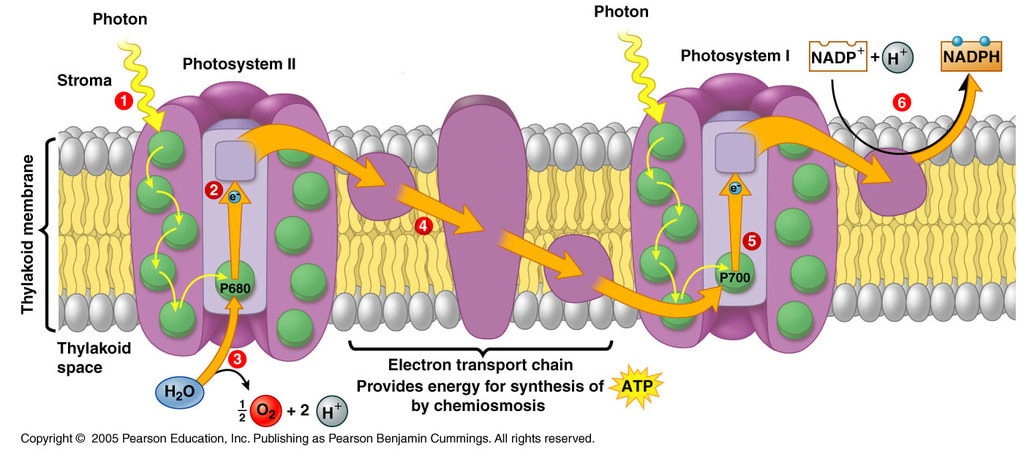
Svetlost apsorbuju dva fotosistema koji apsorbuju svetlost određenih talasnih dužina i to:
- Fotosistem 1 svetlost talasne dužine 700 nanometara
- Fotosistem 2 svetlost talasne dužine 680 nanometara
Kada hlorofil iz hlorofil sistema 1 apsorbuje foton, on prelazi u pobuđeno stanje i ispušta elektron, te se oksiduje.
Otpušteni elektron predaje redox molekulu koji se primanjem elektrona redukuje.
Redox molekul oksidacijom otpušta elektron i predaje ga ferodoksinu koji se redukuje.
Ferodoksin se oksiduje i predaje elektron flavoprotinu. Oksidacijom flavoprotin oslobadja 2 elektrona i predaje NADP 2 vodonikovih jona koji redukuju NADP u NADPH2 (NADPH+H+).
Svi pomenuti molekuli se nazivaju prenosioci elektrona, dok su navedene reakcije oksidoredukcije jer se jedan molekul oksiduje, a drugi redukuje.
Otpušteni elektron predaje redox molekulu koji se primanjem elektrona redukuje.
Redox molekul oksidacijom otpušta elektron i predaje ga ferodoksinu koji se redukuje.
Ferodoksin se oksiduje i predaje elektron flavoprotinu. Oksidacijom flavoprotin oslobadja 2 elektrona i predaje NADP 2 vodonikovih jona koji redukuju NADP u NADPH2 (NADPH+H+).
Svi pomenuti molekuli se nazivaju prenosioci elektrona, dok su navedene reakcije oksidoredukcije jer se jedan molekul oksiduje, a drugi redukuje.
Primanjem fotona iz hlorofil sistema 2 se tako]e oslobađa elektron. Hlorofil se oksiduje i predaje elektron redoks molekulu koji se redukuje. Redox molekul se oksiduje i predaje molekul ferodoksinu. Ferodoksin predaje elektron kveščn molekulu (Q molekulu). Q molekul predaje elektron citohromu f, koji ga potom predaje citohromu b6. Citohrom b6 se oksiduje i vraća molekul hlorofil sistemu 1. Hlorofilsistem 2 svoj elektron nadoknadjuje fotolizom vode ,pri cemu se takodje oslobadja kiseonik. U svetloj fazi voda je glavni donor elektrona, a NADP krajnji akceptor.
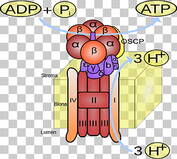
U toku redukcije hlorofil sistema 1 elektroni se premeštaju niz elektrohemijski gradijent prerko serije prenosilaca elektrona. Elektroni se premeštaju iz tilakoida u matrix hloroplasta.Za svaki elektron koji napusti tilakoid ubacuje se jadan H+ proton u lumen tilakoida i na taj način se stvara elektrohemijski gradijent. Odnosno stvara se razlika potencijala izmedju lumena tilakoida i stome (matriksa) hloroplasta. Po zakonu difuzije protoni teže da izađu iz tilakoida kroz membranu , dok je jedino mesto gde mogu to da učine protonska pumpa. Protonska pupa ima ulogu u redukciji adenozin difosfata odnosno sintezi atp. Za svaka 3 h+ jona koja napuste tilakoid sintetiše se jedan atp molekul. Sinteza atp molekula vrši se pod uticajem posebnog enzima ATP sintetaze ili atepeaze koji se nalazi u membrani tilakoida. Ovaj enzim vrši vezivanje neorganskog fosfora za adp prevodeći ga u atp.
Glavni produkti svetle faze fotosinteze su :
- ATP
- NADPH2
- Kiseonik
Proizvodi svetle faze, ATP (sadrži energiju) i NADPH2 (donosi vodonik poreklom iz vode), se koriste u tamnoj fazi da bi se neorganski ugljenik iz CO2 ugradio u organska jedinjenja. To se naziva fiksacija ugljenika i izvodi se u Kalvinovom ciklusu u stromi hloroplasta. Početno jedinjenje ovog ciklusa je istovremeno i završno i naziva se ribulozodifosfat (RuDP). Ugljen-dioksid iz atmosfere ulazi u ćeliju (hloroplaste) i vezuje se za RuDP (ima 5C atoma) pri čemu nastaje jedno nestabilno jedinjenje sa 6C atoma koje se razlaže na dva molekula fosfoglicerinske kiseline (sa po tri С atoma). Da je to prvi stabilan proizvod tamne faze fotosinteze otkrio je Kalvin i za to otkriće 1961. godine dobio Nobelovu nagradu za hemiju. Preko niza proizvoda u toku ciklusa opet se stvara RuDP.Pri okretanju jednog Kalvinovog ciklusa u organska jedinjenja se ugradi jedan C atoma iz CO2. Znači, da bi nastao jedan molekul fruktoze ciklus mora da se okrene 6 puta.
Glavni procesi u kalvinovom ciklusu odnosno tamnoj fazi fotosinteze su:
- Karboksilacija (fiksacija ugljenika)
- Redukcija 3-fosfoglicerinske kiseline
- Regeneracija 1,5 ribuloze bifosfata.
Kalvinov ciklus započinje karboksilacijom 1,5 ribuloze bifosfata. Dobijeno jedinjenje sa 6c atoma je nestabilno i razlaže se na dva molekula fosfoglicerinske kiseline sa po 3C atoma. Uz utrošak ATP-a iz svetle faze 3-fosfoglicerinska kiselina fosforiluje gradeći energetski obogaćenu 1,3 fosfoglicerinsku kiselinu. Pod dejstvom NADPH2 1,3 fosfoglicerinska kiselina se redukuje gradeći 2 izomerne trioze gliceraldehid 3-fosfat (G3P) i dihidroksiaceton-fosfat. Spajanjem ove 2 trioze dobija se šećer sa 6 C atoma fruktoza 1,6 bifosfat. U daljem toku ciklusa dešava se proces konverzije šećera u kome se delovi ugljeničnog lanca sa 6C atoma kombinuju sa triozama i daju šećere sa 4,5,7 C atoma. Konverzijom šećera se stvara ribuloza fosfat koji uz utrošak ATP-a fosforiluje u 1,5 ribulozu bifosfat.
Asimilati, organske materije stvorene u listu transportuju se u sve druge delove tj. organe biljke. Aktivnim transportom se u floem ubacuju asimilati koji se kreću ka svim delovima biljke. U procesu aktivnog transporta se troši ATP odnosno energija.
Faktori koji utiču na fotosintezu su količina svetlosti , količina ugljen dioksida i temperatura.
- Pri konstantnoj temperaturi, brzina ugradnje ugljenika zavisi od svetlosnog zračenja, u početku se ubrzava pri pojačavanju zračenja. Međutim, pri višim nivoima svetlosnog zračenja ova zavisnost više ne važi, a brzina ugradnje ugljenika ostaje konstantna na dostignutom nivou.Razlog tome je što biljka ne može da iskoristi produkte svetle faze koji su se nagomilali, jer se tamna faza odvija sporije.
- Pri konstantnom nivou svetlosnog zračenja, brzina ugradnje ugljenika se pojačava pri porastu temperature u određenom vrlo ograničenom rasponu. Ovaj efekt je primijećen samo pri visokim nivoima svetlosnog zračenja. Pri nižim nivoima, povećanje temperature ima relativno mali uticaj na brzinu ugradnje ugljenika.
- Dok raste koncentracija ugljen-dioksida, brzina kojom se sintetišu šećeri u fazi svetlosno zavisnih reakcija se povećava sve dok je ne ograniče neki drugi faktori.
5. Fotosinteza from ltixomir
